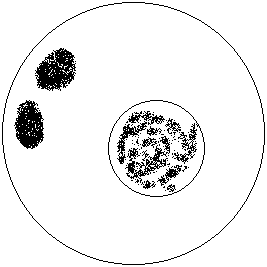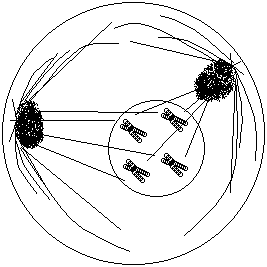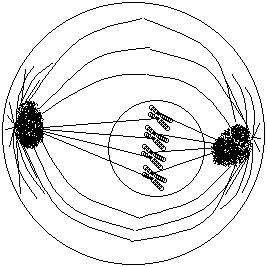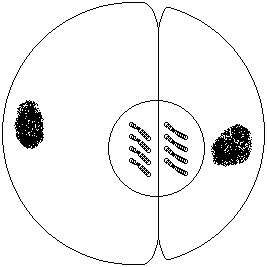
1- سلولی با ژنوتیپ AaBb دارای دو جفت کروموزوم همتا یا 4 کروموزوم است که بر روی هر کروموزوم یکی از آلل ها را دارا می باشد، این سلول توانایی تولید 4 نوع گامت را دارد که در هر تقسیم میوز بسته به آرایش متافازی میوزI فقط دو نوع گامت تولید می کند. با توجه به اینکه در هر آرایش متافازی در میوز I دو نوع گامت تولید می شود(در صورت هتروزیگوت بودن) اگر تعداد انواع گامت ها را بر دو تقسیم کنیم تعداد آرایش متافازی بدست می آید برای بدست آوردن انواع گامت ها هم کافیست انواع گامت های هر صفت را محاسبه کنیم و در هم ضرب کنیم: آرایش متافازی =
برای بدست آوردن انواع گامت های یک صفت به مثال توجه کنید: و انواع گامت هایی که ژنوتیپ های فوق می توانند بسازند برابر است با 4 و 8 که با تقسیم بر دو می توانیم تعداد آرایش های متافازی آنها را بدست آوریم.
فردی با این نوع ژنوتیپ AA Bb DD فقط دو نوع گامت تولید می کند.
کروموزوم های هوموزیگوت پس از جدا شدن تولید 1 نوع گامت می کنند ولی اگر کروموزوم های همولوگ هتروزیگوت باشند پس از جدا شدن از یکدیگر برای هر صفت 2 نوع گامت حاصل می شود. درانسان کروموزوم x نسبت به کروموزووم Y بزرگتر و دارای ژن های بیشتری است پس سلول های پیکری زن سالم از سلول های مرد سالم ژن های بیشتری دارد.
2- در نشانگان ترنر کاریوتیپ xo را نشان می دهد در واقع یک x کم دارد در مبتلایان به نشانگان فریاد گربه در کروموزوم شماره 5 قطعه ای کمبود دارد.
3- هر سلول زاینده انسانی در انجام تقسیم میوز فقط یک نوع آرایش متافازی را می توانند انتخاب کند بنابر این در هر تقسیم میوز فقط 2 نوع گامت حاصل می شود. سلول های انسان با 2n=46 کروموزوم در هنگام تقسیم میوز I تشکیل 23 ساختمان تترادی می دهند و چون هر تتراد (به دلیل متفاوت بودن آلل های کروموزوم های هومولوگ) دو نوع گامت می تواند تولید کند بنابر این سلول های انسانی با 46 کروموزوم توان تولید گامت را دارد.
3- تعداد کل ژنوتیپ ها برابر است با تعداد ژنوتیپ های هموزیگوت به علاوه تعداد ژنوتیپ های هتروزیگوت و با توجه به اینکه تعداد ژنوتیپ های هموزیگوت همیشه برابر تعداد آلل هاست.
کل ژنوتیپ ها = ژنوتیپ هتروزیگوت+ ژنوتیپ هموزیگوت
چون انسان موجودی دیپلوئید است و در هر فرد دو آلل وارد می شود چون الل های فرد هتروزیگوت با هم تفاوت دارند و ترتیب نوشتن آنها هم فرق می کند. تعداد کل ژنوتیپ هتروزیگوت برابر است با ترکیب 2 از مقدار آلل های داده شده. به بیان دیگر از بین 5 نفر به چند طریق می توان کمیته های 2 نفری انتخاب کرد.
4- اگر در جمعیتی برای صفتی n آلل داشته باشیم تعداد کل ژنوتیپ های موجود برابر است با اگر بین آلل ها رابطه غالب و مغلوبی وجود نداشته باشد تعداد کل فنوتیپ ها برابر است با تعداد کل ژنوتیپ ها.
5- اگر تعداد کل آلل ها را بدهند چون آلل ها با ژنوتیپ هموزیگوت برابر است و با توجه به فرمول می توانیم تعداد کل ژنوتیپ ها را مشخص کنیم و با کم کردن از هموزیگوت ها هتروزیگوت ها را بدست آوریم. برای درک بهتر به مثال توجه کنید: اگر 4 آلل داشته باشیم تعداد کل ژنوتیپ ها برابر است با و اگر تعداد آلل ها(که برابر است با هموزیگوت ها) را از تعداد کل کم کنیم هتروزیگوت ها را می توانیم بدست آوریم:
6=4-10
6- اگر در یک شجره نامه فرد ناقل مشاهده شود بنابر این صفت مورد بحث مغلوب است(که در این صورت باید مشخص شود که اتوزومی است یا جنسی). از پدر و مادری که به ظاهر سالم هستند فرزندی که مبتلا به بیماری وابسته به جنس مغلوب است(کور رنگی، هموفیلی، دیستروفی عضلانی و ...) متولد شود این فرزند حتما پسر خواهد بود(یعنی از نوع مذکر و پسر ها هم نیمی سالم و نیمی بیمار خواهند بود) چرا که فرزند دختر باید هر دو کروموزوم x مبتلا را دریافت کند. در این شجره نامه پدر و مادر به ظاهر سالم بوده اند و چون فردی مبتلا به بیماری وابسته به جنس مغلوب به دنیا آمده پس پدر حتما سالم و مادر هم ناقل بوده است در مورد کور رنگی می توانیم این ژنوتیپ ها را برای پدر:XY و این ژنوتیپ را برای مادر متصور شویم: . در بیماری کور رنگی (و کلا بیماریهای وابسته به جنس مغلوب) پسر یا بیمار است یا کاملا سالم ولی دختر به علت داشتن دو کروموزوم XX میتواند ژنوتیپ های سالم XX و ناقل و بیمار را داشته باشد.
7- گروه های خونی: Rh خون توسط دو الل مشخص می شود، افرادی که Rh+ دارند میتوانند دو نوع ژنوتیپ داشته باشند: RR ، Rr و افرادی که Rh- دارند دارای ژنوتیپ rr هستند در واقع Rh سه نوع ژنوتیپ دارد که در دو نوع فنوتیپ خودش را نشان می دهد که عبارتند از: RR , Rr , rr و گروه های خونی A,B, O گروه های خونی دارای 6 نوع ژنوتیپ هستند که در 4 فنوتیپ خود را نشان می دهند.
18=6 نوع ژنوتیپ گروه خونی ×3 نوع ژنوتیپ Rh
8= 4 نوع فنوتیپ گروه خونی× 2 نوع فنوتیپ Rh
8- افرادی که صفت مغلوب را نشان می دهند دارای دو آلل مغلوب هستند که فراوانی هر یک از آنها برابر است. aa=
9- گروه خونی A+ با شرط هتروزیگوت بودن می شود: و هموزیگوت= فرد هتروزیگوت توانایی تولید چهار نوع گامت را داراست. اگر موجودی دارای دو جفت کروموزوم باشد با آلل های مختلف (دو جفت کروموزوم یعنی چهار کروموزوم) هر جفت توانایی تولید دو گامت را داراست و در کل چهار نوع گامت تولید می شود.
10- از آمیزش دو خوکچه ی هندی سیاه هتروزیگوت Bb با هم Bb×Bb روابط زیر را خواهیم داشت:
( B+ b)( B+ b)= BB+ Bb+ bb
با توجه به روابط بالا ژنوتیپ ها هتروزیگوتBb و = × هم هموزیگوت هستند. اگر روابط غالب و مغلوبی وجود داشته باشد آلل غالب با حرف بزرگ نمایش داده می شود.
11- ژنوتیپ ها گروه خونی B+ = , , , و همین طور سایر گروه ها را میتوان نوشت...البته گروه خونی O- به این صورت است:iirr
12- گاهی گروه های خونی را با ژنوتیپ بیمار یها با هم می نویسند برای مثال: فردی که هموفیل است و گروه خونی A+ دارد و برای این صفات هتروزیگوت است می شود: کور رنگی(دالتونیسم) را با نماد c و هموفیل را با نماد h نشان دادیم و چون هتروزیگوت است قادر به تولید حداکثر گامت هاست و 8 نوع گامت می تواند تولید کند(8=2×2×2).
13- پدری که هموفیل است ژنوتیپ آن به این صورت خواهد بود: و ژنوتیپ مادر سالم و هموزیگوتxx است فرزندان این زوج هیچ کدام بیمار نخواهد بود ولی نصف دختران ناقل خواهند شد! به این صورت که:
× xx=( + y)(x)= + xy
14- در بیماری کم خونی داسی شکل با توجه به اتوزومی بودن و مغلوب بودن بیماری ، ژنوتیپ ها اینچنین هستند: (SS,Ss,ss) اگر پدر و مادر سالم و هتروزیگوت باشند بنابراین داریم Ss×Ss
15- در بیمار های اتوزومی مغلوب هیچ گاه نمی توان انتظار داشت از پدر و مادری بیمار فرزندانی سالم یا ناقل بوجود آیند چون در بیمار های اتوزومی فرد بیمار هموزیگوت است و افراد همزیگوت بیمار نمی توانند افراد هموزیگوت سالم یا هتروزیگوت ناقل بوجود آورند چون تنها آلل هایی که بوجود می آید الل های مغلوب است! در بیماری های اتوزومی مغلوب اگر فرد بیمار از پدر و مادر سالم(به ظاهر) بدنیا بیایید پدرو مادر در واقع هر دو ناقل بوده اند((Pp و از آمیزش آنها Pp×Pp چنین نسبت هایی را می توان انتظار داشت: و اگر بخواهیم نسبت دختر ها و پسر ها را بدست آوریم چون این بیماریها اتوزومی هستند و جنسی نیستند باید هر نسبت را در ضرب کرد.
16- در بیماری کور رنگی xx زن سالم و زن ناقل ولی به ظاهر سالم است و زنی مبتلاست و در واقع سه نوع ژنوتیپ داریم که در دو نوع فنوتیپ خود را نشان می دهد.
اگر در شجره نامه پدر و مادر سالم باشند و فرزندانی بیمار وجود داشته باشد آن بیماری مغلوب است چون در صفات وابسته به جنس دختر مبتلا حتما پدری بیمار داشته...
منبع: گاهنامه زیست شناسی